Modes of speciation - organising the mess
So, I have started to consider how the various kinds of speciation relate to each other. The traditional approach is to conceive of speciation as something that happens in sympatry or in allopatry; that is, in the same locale or isolated locales respectively. A third form of speciation is named by Mayr peripatry, in which peripheral populations, almost but not quite detached from the main distribution of the species, evolve independently of the rest like an isolated population would.
Mayr showed allopatric speciation with a diagram that has become iconographic in evolutionary textbooks:
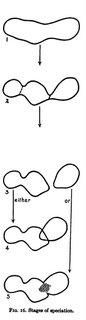
The sequence is to be read from top to bottom over time. The global range of the species, called a metapopulation, fragments as conditions change, perhaps due to geological processes like mountain building or river formation. One part becomes detached, or geographically isolated, it evolves in its own way, incidentally causing reproductive isolation (RI) to evolve. Note that RI is not itself the subject of selection, but happens as a side-effect of other change. This means that being a species formed through RI is not something "functional", but this can change when species that have formed allopatrically come back into contact.
Sympatric populations that are RI will tend to reinforce this isolation through lowered hybrid fitness, since several mechanisms will mean that hybrids actually are less-fit: sexual selection, adaptive traits, chromosomal structure. In the first case, preferential mating may mean that the new-comers find it harder to find willing mates. In the second, the newcomers may be adapted to some resource or condition, while the oldtimers are adapted to a different resource or condition. The hybrids will be neither - sorry - fish nor fowl, and so it will mean they are less viable. In the third case, inversions, translocations, fusions and fissions [see these lecture notes for details] make it harder for the chromosomes of the sex cells to pair up and continue development.
This led Mayr to define species, effectively, as gene pools protected against introgression (the influx of genes from other population):
So, what is the debate? For years now, biologists working in the domain of speciation have argued whether speciation occurs in allopatry, or sympatry or some intermediate peripatry. Much of the debate has been about terms. For example, if the Rhagoletis fruitfly that has speciated by moving from one host (Hawthorns) to another (Apples) is not in sympatry, then nothing is, but it is claimed that the host species is a kind of allopatry. And so on. It's a messy debate, needing some conceptual clarity.
Enter Gavrilets. In the Introduction to his 2004 book Fitness Landscapes and the Origin of Species, he defines these different modes of speciation in ways that - finally - make sense, relating them to each other. In my next post, I will list these and then try to organise them into a "conceptual space".
References
Mayr, Ernst. 1963. Animal species and evolution. Cambridge MA: The Belknap Press of Harvard University Press.
Littlejohn, Murray J. 1969. The systematic significance of isolating mechanisms. In Reflections on systematic biology; Proceedings of an international conference, University of Michigan, June 14Â16, 1967. Washington, DC: National Academy of Sciences.
Mayr showed allopatric speciation with a diagram that has become iconographic in evolutionary textbooks:
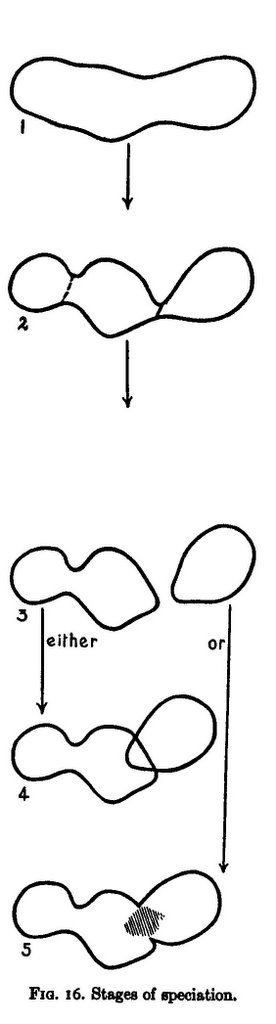
The sequence is to be read from top to bottom over time. The global range of the species, called a metapopulation, fragments as conditions change, perhaps due to geological processes like mountain building or river formation. One part becomes detached, or geographically isolated, it evolves in its own way, incidentally causing reproductive isolation (RI) to evolve. Note that RI is not itself the subject of selection, but happens as a side-effect of other change. This means that being a species formed through RI is not something "functional", but this can change when species that have formed allopatrically come back into contact.
Sympatric populations that are RI will tend to reinforce this isolation through lowered hybrid fitness, since several mechanisms will mean that hybrids actually are less-fit: sexual selection, adaptive traits, chromosomal structure. In the first case, preferential mating may mean that the new-comers find it harder to find willing mates. In the second, the newcomers may be adapted to some resource or condition, while the oldtimers are adapted to a different resource or condition. The hybrids will be neither - sorry - fish nor fowl, and so it will mean they are less viable. In the third case, inversions, translocations, fusions and fissions [see these lecture notes for details] make it harder for the chromosomes of the sex cells to pair up and continue development.
This led Mayr to define species, effectively, as gene pools protected against introgression (the influx of genes from other population):
... species are reproductive communities. The individuals of a species of animals recognize each other as potential mates and seek each other for the purpose of reproduction. A multitude of devices insure intraspecific reproduction in all organisms ... . The species is also an ecological unit that, regardless of the individuals composing it, interacts as a unit with other species with which it shares the environment. The species, finally, is a genetic unit consisting of a large, intercommunicating gene pool, whereas the individual is merely a temporary vessel holding a small portion of the contents of the gene pool for a short time (Mayr, 1963 p21)RI itself became the focus of a lot of research, most recently summarised in Coyne and Orr's book Speciation. RI was divided into subcategories, versions of which found their way into the textbooks. The primary division was between prezygotic and postzygotic or between those mechanisms that prevented the fusion of the sex cells, and those which prevented the fused sex cell, the zygote, from developing through to further reproduction. The most complete list is this one:
A classification of Reproductive Isolating Mechanisms (RIMs)Note that the barriers here are not absolute - the RIMs only produce a reduced frequency of successful breeding.
From (Littlejohn 1969: 461)
1. Reduction of contact
(a) temporal
(b) ecological
2. Reduction of mating frequency
(c) ethological
(d) morphological
PREMATING
----------------------------------------------------------
POSTMATING
3. Reduction of zygote formation
(e) gametic and reproductive tract incompatibilityPREZYGOTIC
----------------------------------------------------------
POSTZYGOTIC
4. Reduction of hybrid survival
(f) hybrid inviability
5. Reduction of gene flow through hybrids
(g) hybrid ethological isolation
(h) hybrid sterility
(i) hybrid breakdown
So, what is the debate? For years now, biologists working in the domain of speciation have argued whether speciation occurs in allopatry, or sympatry or some intermediate peripatry. Much of the debate has been about terms. For example, if the Rhagoletis fruitfly that has speciated by moving from one host (Hawthorns) to another (Apples) is not in sympatry, then nothing is, but it is claimed that the host species is a kind of allopatry. And so on. It's a messy debate, needing some conceptual clarity.
Enter Gavrilets. In the Introduction to his 2004 book Fitness Landscapes and the Origin of Species, he defines these different modes of speciation in ways that - finally - make sense, relating them to each other. In my next post, I will list these and then try to organise them into a "conceptual space".
References
Mayr, Ernst. 1963. Animal species and evolution. Cambridge MA: The Belknap Press of Harvard University Press.
Littlejohn, Murray J. 1969. The systematic significance of isolating mechanisms. In Reflections on systematic biology; Proceedings of an international conference, University of Michigan, June 14Â16, 1967. Washington, DC: National Academy of Sciences.
posted by John S. Wilkins @ 3:41 pm
![]()
![]()





<< Home