The Voltage Gate: Basic Concepts: What Is Ecology?
posted by John S. Wilkins @ 4:06 am
![]()
![]()
Evolution, culture, philosophy and chocolate! John Wilkins' continuing struggle to come to terms with impermanence... "Humanus sum, nihil humanum a me alienum puto" - Terence
posted by John S. Wilkins @ 2:33 pm
![]()
![]()
posted by John S. Wilkins @ 10:23 am
![]()
![]()
posted by John S. Wilkins @ 5:23 pm
![]()
![]()
Tales from the imperfect rural wife: Chuck had it all wrong in the survival gameThe irony comes from the fact that Chuck did actually consider female choice in evolution, in a rather different context, but more from the fact that dandelions have no sex. They are clonal organisms that reproduce from parts if disturbed. Slash them as much as you like, they will keep coming back. Survival of the most stubborn indeed.
by Paula Cassidy
06.02.06
I had just chopped off their heads, but by the next morning, this lawn full of infiltrators resurrected to full attention, awaiting the next bloody battle, standing there semi-headless, taunting me with their “Darwinian survival of the fittest chant.” Chemical warfare crossed my mind, but my heart leans ever so tenderly to the possibility of a happy, healthy planet and Kyoto Protocol and such—unlike our glorious western leaders who live in some confused environmental denial bubble. The low tech war was on. May the most stubborn species win.
History tells of a carpenter that forever changed our lives. Although golf courses and manicured suburban homeowners may deem him the messiah, this carpenter didn’t wander the lands guiding the spiritual walk of the multitudes. Edwin Beard Budding invented the first lawn mower. His 1830’s patent even touts that country gentlemen would not only be amused by his invention, but would also reap the rewards of a little healthy exercise. Country gentlemen. Right then. To the present, where women rule the powerful motorized riding lawn mower, no longer sitting pretty on the sidelines, sipping lemonade in tight corsets and poofy dresses. Vanity mirror? Forget about it. Who wears lipstick to battle?
I roll my engineless reel mower about thirteen times over the same dandelion stem. The mower was a Mother’s Day gift, but before a mob of sympathetic mothers disperse to lynch my husband, it should be noted that it was a gift I had requested; hindsight brought on by frustration, finds me pining for the sapphire ring or a new toaster. No fuel. No noise. No environmental impact. Great cardio workout. The concept was great on paper, but the reality stood before me, relentlessly clinging to life and limb on acres of grass. I roll over the dandelion one more time, and one more time it bends, side stepping its fate. Stubbornness is something we both had in common. It refused death and I refused the effort to bend over and pull it out. Stalemate.
And without warning, like a trumpet sounding in the high afternoon, I hear the call of the machine—the lure of the green John Deere, parked alone and abandoned in the barn, inside the mechanical perimeters of my husband’s fleet of un-environmentally friendly toys and gadgets. Now the dilemma. Stick to my ecological and heart healthy guns, or cave like a hypocritical jellyfish so I can kill me some dandelions real fast-like, and get on with my day. What’s a girl to do?
“You want to use the riding mower, don’t you? Taking too long, eh?” Ah, thank goodness for sarcastic husbands, because without them, how would we women justify our intrinsic stubbornness. Farewell green champion, may you sit idly in the barn, for destiny calls me to the front yard. My clipping shears in hand, I head into battle, the last samurai, facing each adversary one-on-one, with the mutual respect of a true warrior. Snip. Snip. Snip. Our man Charles Darwin got it all wrong, because he failed to consider the female fight for equality. Fittest? No way. Survival of the “stubborn-est” is the best insurance for species predominance.
posted by John S. Wilkins @ 11:18 am
![]()
![]()
posted by John S. Wilkins @ 2:33 pm
![]()
![]()
posted by John S. Wilkins @ 2:27 pm
![]()
![]()
posted by John S. Wilkins @ 10:09 am
![]()
![]()
posted by John S. Wilkins @ 10:00 am
![]()
![]()
Writing this week in the Proceedings of the National Academy of Sciences, scientists say higher temperatures near the equator speed up the metabolisms of the inhabitants, fueling genetic changes that actually lead to the creation of new species.There are many errors in this release, not the least being the definition and explanation of a new species, but one ought not attack scientists for the inability of a journalist (or in this case, PR maven) to express themselves properly.
The finding — by researchers from the University of Florida, the National Center for Ecological Analysis and Synthesis, Harvard University and the University of New Mexico at Albuquerque — helps explain why more living species seem to exist near the equator, a scientific observation made even before naturalist Charles Darwin set sail to South America on the H.M.S. Beagle nearly two centuries ago.
It may also have a bearing on concepts such as global warming and efforts to preserve diversity of life on Earth.
“We’ve shown that there is indeed a higher rate of evolutionary change in the form and structure of plankton in the tropics and that it increases exponentially because of temperature,” said James Gillooly, an assistant professor of zoology with the UF Genetics Institute. “It tells us something about the fundamental mechanisms that shape biodiversity on the planet.”
Speciation — when animals or plants actually evolve into a new species — occurs when life forms with a common ancestor undergo substantial genetic change.
Using a mathematical model based on the body size and temperature-dependence of individual metabolism, the researchers made specific predictions on rates of speciation at the global scale. Then, using fossils and genetic data, they looked at rates of DNA evolution and speciation during a 30-million-year period in foraminifera plankton, a single-celled animal that floats in the ocean. Researchers compared arrivals of new species of this type of plankton with differences in ocean temperatures at different latitudes ranging from the tropics to the arctic. The results agreed closely with predictions of their model.
posted by John S. Wilkins @ 10:47 pm
![]()
![]()
posted by John S. Wilkins @ 2:40 pm
![]()
![]()
A species is a lineage or set of closely related lineages [De Querioz] that clusters genomically [Mallet] through either stochastic [Pie and Weitz] or cohesive [Templeton] mechanisms and processes, which can be due to exogenous selection tracking fitness peaks or endogenous selection for compatibility with genetic exchange, or some admixture of both.Add to this my Synapomorphic Species Concept, which is a general specification of the notion of biological species rather than a particular conception:
A species is a lineage separated from other lineages by causal differences inand you have more than enough from me on species definitions for now...
synapomorphies.
posted by John S. Wilkins @ 2:26 pm
![]()
![]()
The notion that "the" Synthesis was somehow complete at one time or another in its history implies that the participants were aiming at some culminating event, like the Resurrection of Christ.The canonical texts are being treated as if they were The Gospel according to Saint Doby, The Gospel according to Saint Ernst, The Gospel according to Saint G. G., The Gospel according to Saint Julian, The Gospel according to Saint Bernhard, and The Gospel according to Saint Ledyard. Scientists are explorers, not prophets. For them to display themselves otherwise is as dishonest as it is misleading.
Ghiselin, Michael T. (2001), "Evolutionary synthesis from a cosmopolitan point of view: a commentary on the views of Reif, Junker and Hossfeld", Theory in Biosciences 120:166-172.Can I get an amen!? Amen, brother.
posted by John S. Wilkins @ 1:47 pm
![]()
![]()
James Phillips, professor of anthropology at the University of Illinois at Chicago, said that it was wrong to suggest that the stone tools could have been made by earlier species of humans, such as Homo erectus, a creature that evolved more than 1.8 million years ago and predated modern humans by many hundreds of thousands of years.Good to see this line being taken. If the evidence suggests they did make stone tools, then intuitive preconceptions about brain size and tool making have to go by the wayside. As Grissom says, the evidence does not lie.
"These tools are so advanced that there is no way they were made by anyone other than Homo sapiens," Professor Phillips said.
Now, however, another team of stone-tool experts has cast doubt on this judgement, saying that similar stone tools have been uncovered on the island that clearly predate the arrival of modern Homo sapiens.
Adam Brumm of the Australian National University in Canberra and his colleagues report in the journal Nature that they have found hundreds of almost identical stone tools at a site called Mata Menge just 30 miles away from the Liang Bua cave. They say the tools are between 700,000 and 840,000 years old - too old to have been made by Homo sapiens - and that the production techniques are practically identical to that used at Liang Bua 18,000 years ago.
posted by John S. Wilkins @ 12:35 pm
![]()
![]()
 Israeli researchers have described eight new species of crustaceans and invertebrates in a recently discovered limestone cave isolated from the external world. They live in and around an underground lake fed by deep water sources rather than rainfall from above.
Israeli researchers have described eight new species of crustaceans and invertebrates in a recently discovered limestone cave isolated from the external world. They live in and around an underground lake fed by deep water sources rather than rainfall from above.
posted by John S. Wilkins @ 10:52 am
![]()
![]()
posted by John S. Wilkins @ 10:40 am
![]()
![]()
that have pretended to find the truth of all natural philosophy in the Scriptures; scandalizing and traducing all other philosophy as heathenish and profane. But there is no such enmity between God's word and His works; neither do they give honour to the Scriptures, as they suppose, but much imbase them. For to seek heaven and earth in the word of God, (whereof it is said, Heaven and earth shall pass, but my word shall not pass,) is to seek temporary things amongst eternal: and as to seek divinity in philosophy is to seek the living amongst the dead, so to seek philosophy in divinity is to seek the dead amongst the living: neither are the pots or lavers, whose place was in the outward part of the temple, to be sought in the holiest place of all, where the ark of the testimony was seated. And again, the scope or purpose of the spirit of God is not to express matters of nature in the Scriptures, otherwise than in passage, and for application to man's capacity, and to matters moral or divine.
Advancement of Learning, Bk II.XXV.16, p216-7 in the Everyman edition.
I'll get back to microbial species, I promise...
posted by John S. Wilkins @ 2:47 pm
![]()
![]()

posted by John S. Wilkins @ 1:39 pm
![]()
![]()
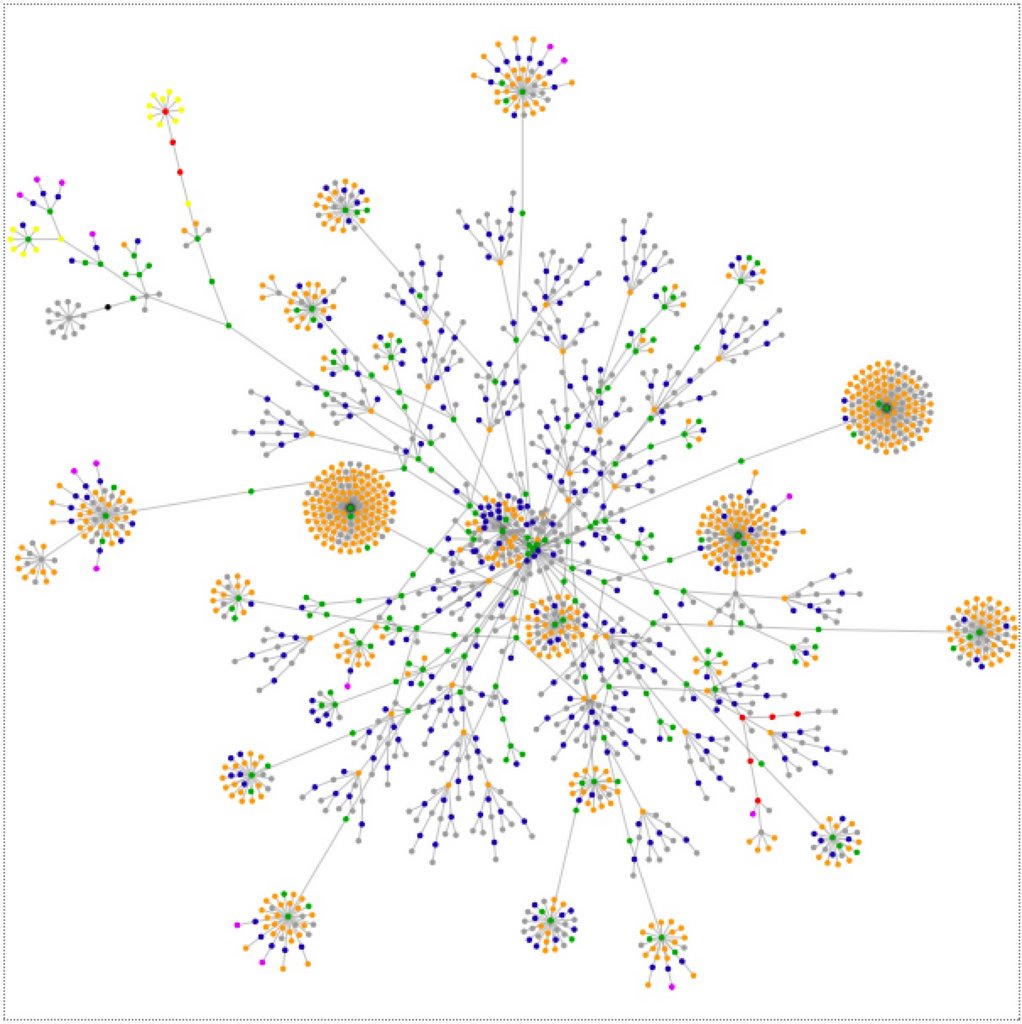 Well, everybody's doing it (doing it, doing it) so I have to. Yes, if everybody else jumped off a cliff I would too, mum. This is what the HTML tags look like for this site when run through the websitesasgraphs Java Applet. It is also what the mental contents of my my brain look like when viewed through any medium at all.
Well, everybody's doing it (doing it, doing it) so I have to. Yes, if everybody else jumped off a cliff I would too, mum. This is what the HTML tags look like for this site when run through the websitesasgraphs Java Applet. It is also what the mental contents of my my brain look like when viewed through any medium at all.
posted by John S. Wilkins @ 1:32 pm
![]()
![]()
posted by John S. Wilkins @ 10:50 am
![]()
![]()
posted by John S. Wilkins @ 8:45 pm
![]()
![]()


posted by John S. Wilkins @ 10:30 pm
![]()
![]()

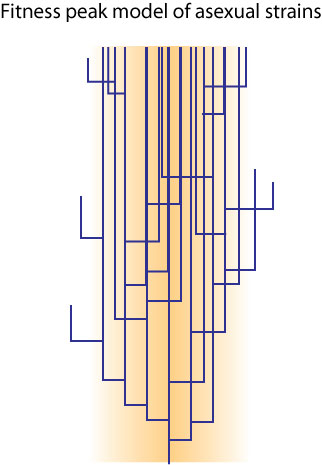
posted by John S. Wilkins @ 3:15 pm
![]()
![]()
 I mentioned prokaryotes before. This refers broadly to a paraphyletic group of organisms that are basically not-eukaryotes. Nowadays we refer instead to several groups: Bacteria, Archaea (which is sometimes decomposed into several other groups), and Eukaryota. One thing the not-Eukaryotes have in common, though, is a lack of a nuclear membrane, allowing the transfer of genetic material between genomes.
I mentioned prokaryotes before. This refers broadly to a paraphyletic group of organisms that are basically not-eukaryotes. Nowadays we refer instead to several groups: Bacteria, Archaea (which is sometimes decomposed into several other groups), and Eukaryota. One thing the not-Eukaryotes have in common, though, is a lack of a nuclear membrane, allowing the transfer of genetic material between genomes.
 I failed to mention a process that is directly analogous to sex in bacteria - conjugation. This is a case where part of the genetic component, usually plasmids, which are secondary small chromosomes (B), or the main nucleoid (A), can be inserted into another cell via processes called pili, which are part of the Type IV secretory system used for other purposes and which is homologous to flagella. The typical mode of conjugation is that one mating type (often called the "male") is activated by pheromones from another mating type ("female") to attach the pilus to the recipent, and insert the genetic material.
I failed to mention a process that is directly analogous to sex in bacteria - conjugation. This is a case where part of the genetic component, usually plasmids, which are secondary small chromosomes (B), or the main nucleoid (A), can be inserted into another cell via processes called pili, which are part of the Type IV secretory system used for other purposes and which is homologous to flagella. The typical mode of conjugation is that one mating type (often called the "male") is activated by pheromones from another mating type ("female") to attach the pilus to the recipent, and insert the genetic material.
posted by John S. Wilkins @ 11:07 am
![]()
![]()

posted by John S. Wilkins @ 5:19 pm
![]()
![]()
The title of this paper is slightly modified from that of an article I encountered some years ago, which appeared to be approaching the problem of the relationships of the Lagomorpha, or rabbits and their relatives, from the most basic point of view. This paper, entitled “Gibt es Leporiden?”, seemed to be questioning the very existence of such animals. Investigation showed, however, that the question involved was not whether members of the family Leporidae existed, but whether rabbit-hare hybrids did. Since then, I have met no one who questions the existence of rabbits and hares, and I have been reluctantly forced to accept them.They don't write papers like that any more. I, too, question the reality of rabbits. Just because.
posted by John S. Wilkins @ 12:24 pm
![]()
![]()
posted by John S. Wilkins @ 11:40 am
![]()
![]()
posted by John S. Wilkins @ 1:42 pm
![]()
![]()
posted by John S. Wilkins @ 1:14 pm
![]()
![]()
Fundamentalism
The belief that basic elements of play - like passing, ball handling, and defense - are the essential building blocks of a winning basketball team is generally referred to as "fundamentalism." The fundamentalists formulated their doctrine in the 1980s against the showy, heretical play of Magic Johnson's Los Angeles Lakers. Leading fundamentalist institutions include Bob Jones University and Syracuse. Larry Brown's failure to get the Knicks into the playoffs has been seen as a major setback for the cause of fundamentalism.
posted by John S. Wilkins @ 11:21 am
![]()
![]()
posted by John S. Wilkins @ 6:36 am
![]()
![]()
posted by John S. Wilkins @ 10:08 pm
![]()
![]()
posted by John S. Wilkins @ 1:18 pm
![]()
![]()
posted by John S. Wilkins @ 9:50 am
![]()
![]()
As a general rule, wild animals are at their wildest in Asia, at their boldest in Europe, and most diverse in form in Libya; in fact, there is an old saying, ‘Always something fresh in Libya.’"Libya" is the name for Africa, from the Hellene perspective, of course. Aristotle had another long standing account of biodiversity too - the effects of local water, soil and climate. This view wasn't abandoned until the early years of the 20th century. But it's hybrids that fascinate me and motivate this post.
It would appear that in that country animals of diverse species meet, on account of the rainless climate, at the watering-places, and there pair together; and that such pairs will often breed if they be nearly of the same size and have periods of gestation of the same length. For it is said that they are tamed down in their behaviour towards each other by extremity of thirst.
posted by John S. Wilkins @ 9:04 am
![]()
![]()
posted by John S. Wilkins @ 8:05 pm
![]()
![]()
posted by John S. Wilkins @ 10:16 am
![]()
![]()
posted by John S. Wilkins @ 4:11 pm
![]()
![]()
posted by John S. Wilkins @ 12:27 pm
![]()
![]()

My main blog is Evolving Thoughts.




