Modes of speciation 2: flow versus place
[I'm sorry this has been delayed. I moved house...]
Gavrilets notes that one can use any of the isolating processes to classify speciation, but that the traditional way is to frame it in terms of migration between diverging populations (which is to say, it's what Mayr said in 1942). He lists the varieties I mentioned last time: allopatric, sympatric and parapatric, and aligns them along a single axis of migration rate:
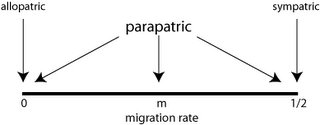
Of interest here is that some of the standard forms of speciation don't appear. For example, speciation by chromosomal rearrangement ("stasipatry", defined by Michael White in the 1970s) is missing. It is clearly in sympatry, but there is no "migration" between the new population and the old (except in cases like the Sphalax ehrenberghi mole rat in Israel, which has major chromosomal races, some of which are mutually infertile, but there is introgression between them). Polyploidy - or the duplication of entire chromosomal complements, common in plants and arguably in animals - is also missing.
Gavrilets deals with them by calling them "scenarios" and "patterns" of speciation. He lists the following cases
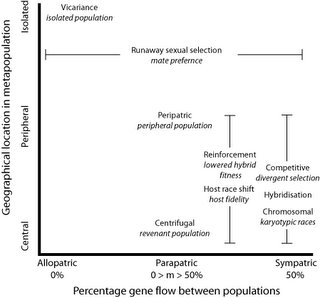
Replacing percentage of gene flow for migration rate and geography as the other axis, we now get a clear picture - sympatry-allopatry is not the only important difference between these "scenarios" or modes of speciation. So also is the relation of the populations to each other. I have placed the sexual selection example between peripheral and isolated because I doubt it would occur species-wide, and is therefore less likely to be a central population case, but it could occur in revenant populations.
We are left with two groups of modes - reinforcement and host race shift (in the case of almost-sympatric parapatry: again it need not be so specific), and competitive, hybrid and karyotypic. Each of these is distinguished by a particular mode or mechanism of speciation.
In my next post I will explore some of the implications of this for the speciation debate, and also for the definitions of "species" used by biologists.
Gavrilets notes that one can use any of the isolating processes to classify speciation, but that the traditional way is to frame it in terms of migration between diverging populations (which is to say, it's what Mayr said in 1942). He lists the varieties I mentioned last time: allopatric, sympatric and parapatric, and aligns them along a single axis of migration rate:

Of interest here is that some of the standard forms of speciation don't appear. For example, speciation by chromosomal rearrangement ("stasipatry", defined by Michael White in the 1970s) is missing. It is clearly in sympatry, but there is no "migration" between the new population and the old (except in cases like the Sphalax ehrenberghi mole rat in Israel, which has major chromosomal races, some of which are mutually infertile, but there is introgression between them). Polyploidy - or the duplication of entire chromosomal complements, common in plants and arguably in animals - is also missing.
Gavrilets deals with them by calling them "scenarios" and "patterns" of speciation. He lists the following cases
- Vicariant - divergent selection and stochastic factors like drift after division of a population by extrinsic factors such as geographical changes;
- Peripatric - a small subpopulation, mostly isolated, at the extreme of the parent range. The idea is that it will have both a non-standard sampling of alleles, and also be subjected to divergent selection pressures in extreme environments (for that species);
- Centrifugal - central populations that carry a sample of many alleles which become isolated through, say, "island" formation (such as the mountain "islands" in the Amazon);
- punctuated equilibrium - the appearance of relatively rapid speciation and subsequent stasis as the population reaches equilibria of alleles. This is a "pattern" rather than a "process" as such, and can be caused by any of the other scenarios;
- Chromosomal speciation;
- Hybridisation - including allopolyploidy. In this case the genetic complement of two species is paired up by a loss of secondary division, giving a symmetrical set of chromosomal strands;
- Reinforcement - once hybrids are of lowered fitness for whatever reason, selection will tend to reinforce separation of the gene pools (for example, a hybrid rock and grass dwelling lizard might be less able to survive in either environment as well as the "pure" lines);
- Competitive - this is Darwin's scenario. Members of a species that are in strong selection for a limited resource may result in specialised forms that are thus in less competition with the ancestral forms that make use of some other resource;
- Clinal - he calls it "speciation along environmental gradients", where limited migration and selection leads to aggregation of forms at the terminal ends of the cline;
- Host shift - this is the famous case of the Rhagoletis fruit flies I mentioned, that Stuart Berlocher and colleagues have studied. Host fidelity replaces geographic isolation;
- Runaway sexual selection - this is obvious.
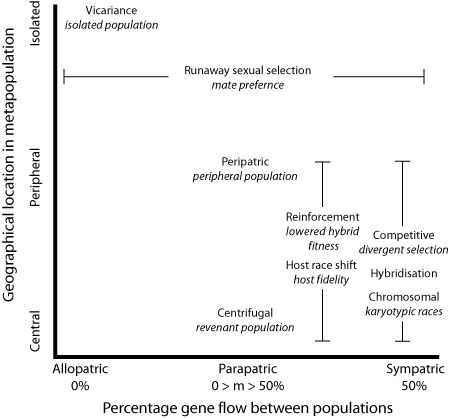
Replacing percentage of gene flow for migration rate and geography as the other axis, we now get a clear picture - sympatry-allopatry is not the only important difference between these "scenarios" or modes of speciation. So also is the relation of the populations to each other. I have placed the sexual selection example between peripheral and isolated because I doubt it would occur species-wide, and is therefore less likely to be a central population case, but it could occur in revenant populations.
We are left with two groups of modes - reinforcement and host race shift (in the case of almost-sympatric parapatry: again it need not be so specific), and competitive, hybrid and karyotypic. Each of these is distinguished by a particular mode or mechanism of speciation.
In my next post I will explore some of the implications of this for the speciation debate, and also for the definitions of "species" used by biologists.
posted by John S. Wilkins @ 1:04 pm
![]()
![]()





<< Home