More from the nether regions of my cortex on the work and ideas of
Sergey Gavrilets...
The idea that there are neutral networks - that is, ridges of nearly identical fitness value in the space of all possible genotypes for a species - suggests a way of reconciling two concepts that have been held to be opposed in the
Darwin Wars: adaptation and drift.
This goes back to a debate between
R. A. Fisher, who first successfully argued that Mendel and Darwin could be reconciled, and who proposed the "fundamental theorem" of selection (which is not a theorem), and
Sewall Wright, who suggested that Fisher's theorem only applied to unrealistic populations (that were infinite and equally likely to mate between any two individuals), and that evolution in fact relied upon changes due solely to statistical or stochastic effects of sampling error in small populations. Wright, of course, is the inventor of the
adaptive landscape. Fisher influenced the British evolutionists, while Wright influenced Dobzhansky and Mayr in America.
Mayr's account of speciation relied on the small population stochasticity of peripheral isolate populations of a species being subjected to fluctuations unrelated to selection for reproductive isolation. In effect, reproductive isolation was a side-effect of evolution in small populations. The British tradition of Haldane and Maynard Smith tended to ignore speciation and focus mostly on selection.
In Gavrilets' view, an adaptive landscape is almost always going to have roughly equivalent regions that are interconnected - that is, they form networks. But they are not selectively neutral networks - each point on the network pathway must be roughly the same fitness. In short, the networks are fitness
ridges or
contours along which species or populations will transcribe a "random walk".
Yep, that's right -
both selection and drift are in play here. The fitness is maintained, but the populations will take random directions due to stochastic effects, for any realistically sized population. So speciation may be due to stochastic effects while selection maintains the
current genotype at the local optimum. That is, in other words, the fitness isn't a
peak, it's a
ridge which is part of a series of interconnected ridges. Selection can only maximise local fitness, not where on the fitness ridge the population is.
Gavrilets uses a different metaphor, and I must point out that it is indeed a metaphor not a model, of a
holey landscape, in which there is a high region of fitness in the landscape, interspersed with regions of low fitness (see figure). In the high fitness region, a random walk can take you all over the place. Of course, one thing Gavrilets insists upon is the high dimensionality of the space of fitness combinations - the 2-dimensional space here is a necessary limitation of paper.
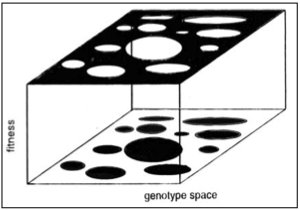
So, what does this mean for speciation? More on this later. For now, I find this enormously interesting: that selection and drift are, in effect, decoupled. They aren't antonyms, they are just different processes.
That makes a lot of the argument between adaptationists and antiadaptationists otiose, I think.
More later









