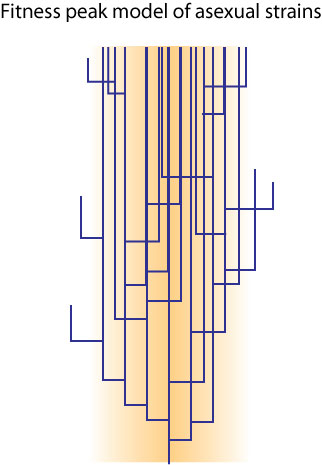
The cluster of genomes of asexual organisms forms what is called a "phylotype" (Denniston 1974, a term coined by C. W. Cotterman in unpublished notes dated 1960; I like to track these things down). Phylotype is a taxon-neutral term, though, that is determined entirely by the arbitrary level of genetic identity chosen. For example, "species" in asexuals might be specified as being 98%+ similarity of genome, or it might be 99.9%+ (I have seen both in the literature). A phylotype of, say 67% or 80% might be used for other purposes (such as identifying a disease-causing group of microbes).
The phylotype concept, while useful in other respects, reinvents (or preinvents - 1960 predates the work of Sokal and Sneath) phenetics. A phenetic taxon was called the Operational Taxonomic Unit (OTU) and it used an arbitrary measure of similarity and difference: an 80% "phenon line" was the arbitrary measure for species based on phenotypic similarities. The problem was that the distances changes as you chose different principal components, and I warrant the same is true for phylotypes.
One solution to the Problem of Homogeneity for asexuals is what I will call the Recombination Species Concept. Proposed by Dykhuizen and Green in 1991, it basically the Biological Species Concept for lineages that occasionally share genes or gene fragments through lateral transfer.
There are several mechanisms by which lateral transfer of DNA can occur among "prokaryotes". One is
DNA fragment reuptake, in which DNA from a cell that has lysed (its membrane or wall has disintegrated, releasing the cell contents into the medium) is taken up by another cell, and rather than being digested it becomes active. There are variations on this. Entire chromosomal rings, called
plasmids can be taken up this way. Or a cell can "bleb", forming vesicles or compartments of lipids, containing DNA (including plasmids), which then attach to the receiving cell, opening up to the interior.

I mentioned prokaryotes before. This refers broadly to a paraphyletic group of organisms that are basically not-eukaryotes. Nowadays we refer instead to
several groups: Bacteria, Archaea (which is sometimes decomposed into
several other groups), and Eukaryota. One thing the not-Eukaryotes have in common, though, is a lack of a nuclear membrane, allowing the transfer of genetic material between genomes.

So the Recombination model of microbial species is based on the claim that the greater the genetic distance between strains, the less likely it is that the genes will be functional and useful in the receiving strain, and this is what serves to maintain the homogeneity of bacterial and other microbial species. One of the claims is in fact that the differences in genetic structure, and in some cases differences in restriction enzymes, that break DNA sequences, will make a nonsense of the DNA fragment or insert it in a nonfunctional location in the target genome.
These compatibility issues act like sex does to maintain the overall "location" in genome space of the population. Those strains that deviate too far from the mode will be unable to take up the functionally useful lateral genes, and so will be more susceptible to extinction through genetic load.
So the Recombination model is a mix of
Maynard Smith's theory of sex, and Mayr's notion of biological species. The only problem is that it isn't consistently true. A beautiful explanation is spoiled by ugly facts. Recombination via lateral transfer appears to be rather more profligate than first appeared (Beiko,
et al. 2005), and there are some "species" of bacteria, such as the Lyme Disease-causing spirochete
Borrelia burgdorferi, that appear not to share genes much, if at all (as Dykhuizen himself observes, Dykhuizen and Baranton 2001). So if in those cases clustering occurs, it is not due to lateral transfer, but some other processes.

I failed to mention a process that is directly analogous to sex in bacteria - conjugation. This is a case where part of the genetic component, usually plasmids, which are secondary small chromosomes (
B), or the main nucleoid (
A), can be inserted into another cell via processes called
pili, which are part of the Type IV secretory system used for other purposes and which is homologous to flagella. The typical mode of conjugation is that one mating type (often called the "male") is activated by pheromones from another mating type ("female") to attach the pilus to the recipent, and insert the genetic material.
Now this is "almost-sex", because there is no genetic reassortment, but other processes will tend to shuffle genes into the nucleoid, as well as utilising the taken-up plasmids. But again, while the mating types seem to act as cohesive mechanisms, conjugation can be profilgate across large, even vast, phylogenetic distances (and hence genetic distances). It has been observed between bacteria and yeast, bacteria and plants, and there has even been a case in which it was observed between bacteria (
E. coli) and mammalian (hamster) cells (Waters 2001).
So while it may be that recombination of lineages through partial genetic transfer operates as a reason for
some phylotypes, it does not account for all, and is therefore not a
sine qua non of specieshood, or homogeneity, among microbes.
Next, we'll consider ecological accounts, as well as drift, migration and geographical isolation.
Beiko, Robert G., Timothy J. Harlow, and Mark A. Ragan (2005), "Highways of gene sharing in prokaryotes", Proceedings of the National Academy of Sciences, USA 102 (40):14332-14337.
Denniston, Carter (1974), "An extension of the probability approach to genetic relationships: One locus", Theoretical Population Biology 6 (1):58-75.
Dykhuizen, D. E., and G. Baranton (2001), "The implications of a low rate of horizontal transfer in Borrelia", Trends in Microbiology 9 (7):344-350.
Dykhuizen, D. E., and L. Green (1991), "Recombination in Escherichia coli and the definition of biological species", Journal of Bacteriology 173 (22):7257-7268.
Waters, V. L. (2001), "Conjugation between bacterial and mammalian cells", Nature Genetics 29 (4):375-376.